
Jul 11, 2026 · 17 min read · abiogenesis
The Spark: How Chemistry Learned to Live
Abiogenesis is the biggest open problem in biology, and the last few years have been some of the field's best. Asteroid samples, the handedness of your own molecules, protocells nobody designed, and a short list of nearby worlds where it might have happened twice.
 A planet still cooling off from its own birth
A planet still cooling off from its own birth
Somewhere between 4.4 and 3.7 billion years ago, on a planet still cooling off from its own birth, a batch of ordinary chemistry did something chemistry had never done. It made a copy of itself. Then the copy made copies. And somewhere in that loop, matter started, in some faint and stubborn way, to keep score.
We don't know how it happened. That's the honest headline. Abiogenesis, the origin of life from non-life, is the biggest open problem in biology, and it has shrugged off a clean answer through seventy years of serious work.
But "unsolved" undersells what's been going on lately. The last couple of years have been some of the field's best, and the interesting part is why. The answer keeps turning out to be interdisciplinary in ways nobody planned for. To chase it you end up reading asteroid geochemistry one week and particle physics the next, with detours through thermodynamics and information theory. Here's the shape of the mystery, and where it's actually moving.
The grocery list came from space
Start with what should be the hard part and turns out to be almost embarrassingly easy: the ingredients.
In 1952, Stanley Miller ran a spark through a flask of simple gases and water and made amino acids. Big moment, and for decades the takeaway was "life's building blocks are chemically cheap." True enough. But the real jolt came from looking up.
In September 2023, NASA's OSIRIS-REx mission dropped a capsule into the Utah desert carrying about 120 grams of the asteroid Bennu, a carbon-rich rock that had been drifting essentially untouched for 4.5 billion years. When labs opened it, a prebiotic starter kit was just sitting there in the dust. Fourteen of the twenty amino acids life on Earth uses to build proteins. All five of the nucleobases that spell out DNA and RNA: adenine, guanine, cytosine, thymine, uracil. Ammonia, in surprising amounts. Formaldehyde, which reacts with ammonia to make yet more amino acids. Phosphates, the backbone material of nucleic acids. And sodium-rich salts, the mineral signature of ancient brine: evaporated pools of alkaline water on the asteroid's long-dead parent body. A chemistry lab, in other words, and the water it ran in.
 Material returned from asteroid Bennu by NASA's OSIRIS-REx mission
Material returned from asteroid Bennu by NASA's OSIRIS-REx mission
Then in late 2025 a fresh analysis of the same sample turned up tryptophan. Tryptophan is one of the trickier amino acids, and it had never been seen in any meteorite or returned sample before. Finding it in pristine asteroid material is a strong sign this chemistry is real, and not Earthly contamination sneaking into the tubes.
Bennu isn't a fluke. Japan's Hayabusa2 brought back grains of the asteroid Ryugu with amino acids in them. The Murchison meteorite, which fell in Australia in 1969, has been a piñata of extraterrestrial organics for half a century. Nucleobase precursors have even been caught forming in the ices of interstellar space.
Which means the ingredients were never the mystery. The recipe is.
The asymmetry that shouldn't be there
Before the recipe, we have to talk about the strangest clue in the case, and the one I find hardest to put down: handedness.
Most of life's important molecules are chiral. Like your hands, they come in two mirror-image versions you can't lay on top of each other, a left-handed (L) form and a right-handed (D) form. The two are chemically identical in nearly every respect. Synthesize amino acids in a beaker with ordinary chemistry and you get a clean 50/50 mix. Flip a coin a billion times, get roughly half heads.
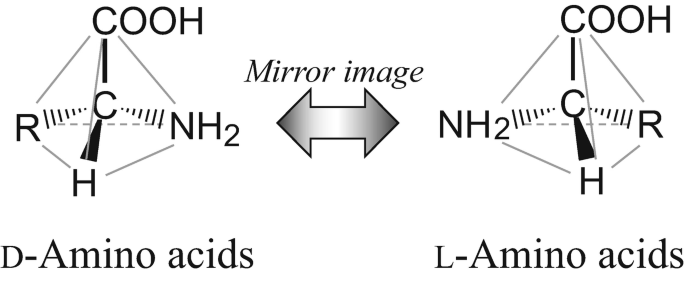 The two mirror-image forms of an amino acid: left-handed (L) and right-handed (D)
The two mirror-image forms of an amino acid: left-handed (L) and right-handed (D)
Life doesn't play the odds. It builds its proteins almost entirely from left-handed amino acids, and its nucleic acids from right-handed sugars. No exceptions worth mentioning: not across the roughly 8 million species alive today, not in anything extinct we've been able to check, all the way down to the root of the tree. Biologists call this homochirality, and where it came from is a real blank.
Why care? Because it's a frozen accident that got locked in at the very start and never thawed. A protein made of mixed left and right amino acids can't fold into anything useful. A strand of RNA with mixed-handed sugars can't zip into a stable helix. So homochirality isn't a cosmetic detail. It may be a precondition for the machinery to work at all. Whatever happened at the origin either needed a chiral bias or manufactured one, and we'd very much like to know which.
This is where it gets strange, because the leading explanations reach all the way down into physics.
Take the space angle first. Stars form inside enormous clouds where light gets scattered and filtered until some of it comes out circularly polarized, corkscrewing with a fixed twist. Circularly polarized UV light treats left- and right-handed molecules differently: it tends to chew up one hand and spare the other. Shine it on a racemic soup long enough and you're left with a slight surplus of one form. Astronomers found exactly this kind of circular polarization in the Orion star-forming region back in 1998, and a 2025 survey from the National Astronomical Observatory of Japan reported that it seems to be a universal feature of regions where stars and planets are being born. The picture is seductive. The twist of light around a newborn star biases the amino acids frozen in nearby ice, and those pre-biased molecules hitch a ride down to Earth with the imbalance already baked in.
For a while, Murchison seemed to back this up. Some of its amino acids showed a left-handed excess, anywhere from a few percent up to fifteen, and pointing in the same direction Earth life eventually chose.
As of 2026, the whole question is cracked back open. If the bias didn't ride in from space, then homochirality got bootstrapped here, probably through some runaway process where one hand, once slightly ahead, amplifies its own lead and quietly poisons its mirror twin.
And underneath all of it sits a whisper from particle physics. The weak nuclear force is the only force we know of that isn't mirror-symmetric. It violates parity. At the most fundamental level, the universe can tell left from right, and that shows up as a real, calculable energy difference between a molecule and its reflection. The catch is that it's minuscule, something like one part in 10^17. Almost certainly too small to have decided anything. But "almost certainly" is carrying a lot of weight in that sentence, and the idea that the handedness of your proteins might be a giant, four-billion-year echo of a broken symmetry in the Standard Model is the sort of thing that keeps people awake.
So the twist in your own molecules might trace back to the light of dying stars, or to a symmetry that breaks at the bottom of physics, or to some accident on the early Earth that never got a second look. We don't know which. We don't even know whether it's one story or three.
The recipe: from a pile of parts to a thing that lives
Now the genuinely hard part. Having amino acids and nucleobases lying around is like owning a warehouse full of engine parts. It is not a car, and it will not turn into one by sitting there.
Life needs a few things a heap of molecules doesn't have. A way to store information. A way to copy it. A way to catalyze its own chemistry. And a boundary to hold the whole racket together. Try to line those up and you smack straight into the field's oldest headache, the chicken-and-egg problem. In a modern cell, DNA holds the instructions, but reading and copying DNA takes protein enzymes, and those enzymes can only be built from the instructions in the DNA. Each needs the other. Neither works alone. So which showed up first?
The RNA world is the most influential answer, and it's a lovely piece of reasoning. RNA is a molecular double agent. Like DNA, it can carry a sequence. Unlike DNA, it can also fold into shapes that catalyze reactions. Those catalytic RNAs are called ribozymes, and pinning them down won a Nobel Prize. If early life ran on RNA doing both jobs, playing recipe and cook at once, the chicken-and-egg knot just comes undone. And there's a fossil of that era hiding in plain sight. The ribosome, the machine every cell uses to assemble proteins, has a catalytic heart made of RNA, not protein. To this day, life builds its proteins using a ribozyme.
The RNA world has its own troubles, mind you. RNA is fragile. Coaxing it to assemble from scratch and then copy itself without enzymes has been miserably hard to pull off in a lab. Which is why the newest thinking has quietly shifted the frame.
Systems chemistry drops the question "which molecule came first" for "which network came first." The insight, pushed hardest by John Sutherland, is that the building blocks needn't arrive one at a time down separate assembly lines. Start with something dead simple and abundant, like hydrogen cyanide, and under plausible early-Earth conditions you can get amino acids, nucleotide precursors, and lipids emerging out of the same chemistry, together. Maybe life wasn't assembled part by part. Maybe it precipitated out of a coupled reaction network more or less at once, as a system.
A 2025 experiment made this weirdly vivid. Researchers reran the old Miller-type spark setup and finally noticed something everybody had looked past for seventy years: the glassware was doing chemistry. Silica in the borosilicate reactors nudged a thin film of HCN polymer to form at the water-gas interface, and out of that film, hollow cell-like blobs, protocells, budded off on their own. So the same rig that cooked the organic "software" was also, in the same spot at the same time, spitting out compartment "hardware." Membranes and molecules, showing up together. Nobody designed it that way. It had probably been happening in those flasks since the 1950s while everyone stared at the amino acids and missed the bubbles.
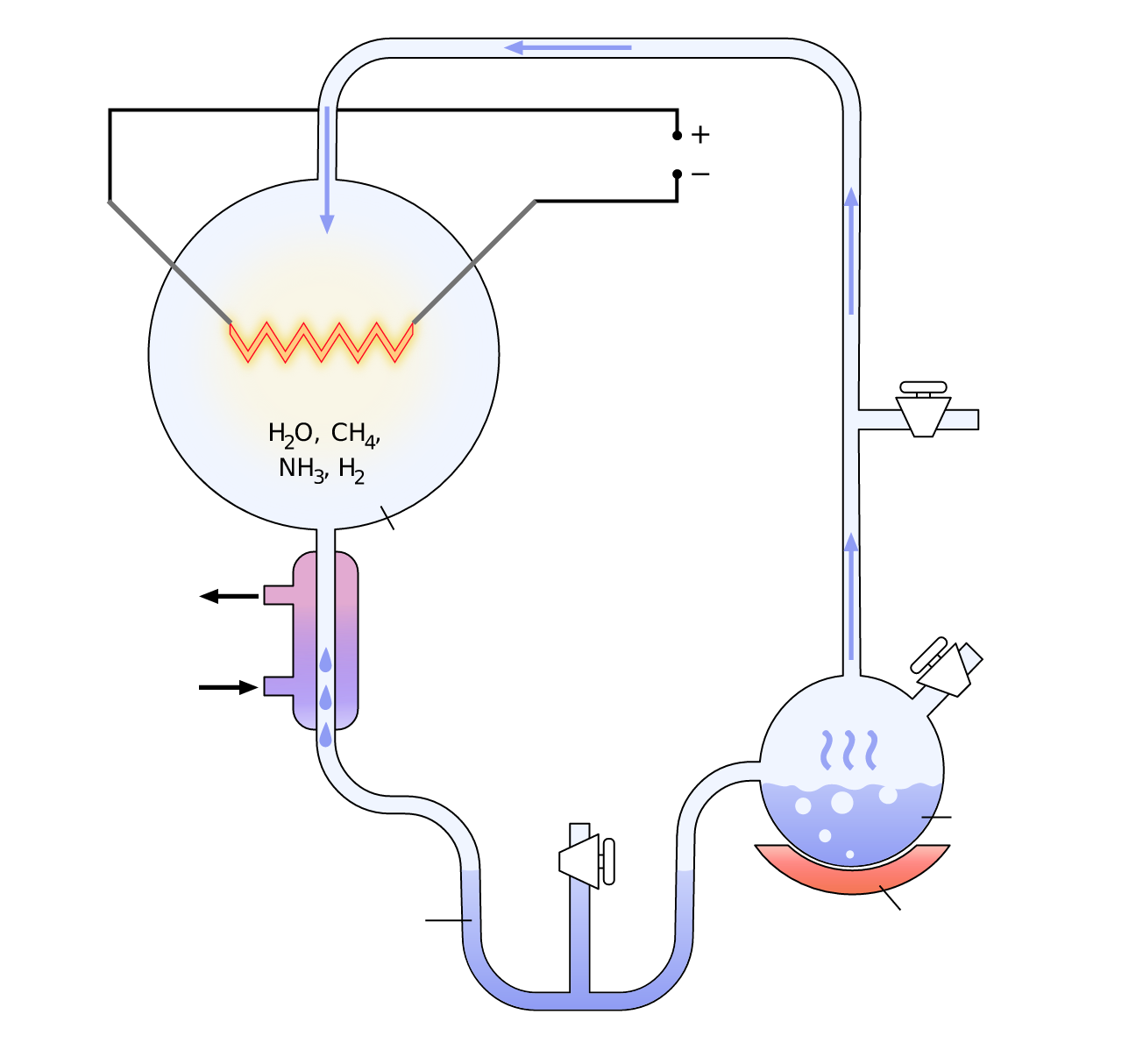 The Miller-Urey apparatus: gases, water, and an electric spark
The Miller-Urey apparatus: gases, water, and an electric spark
There's a related idea that the first compartments didn't even need membranes. Coacervates are oily droplets that form spontaneously and behave like membraneless cells. They pull RNA and its reaction partners into a dense little pocket, where chemistry that would be hopelessly dilute out in the open ocean suddenly finds itself crowded enough to work. A cell before there was a cell.
Where did it happen, and how far back does the trail go?
Two landscapes hog the argument.
Hydrothermal vents come first. Down on the deep seafloor, mineral-rich water seeps through the crust and reacts with rock in a process called serpentinization, which pumps out hydrogen gas and grows tall chimneys shot through with tiny mineral pores. Those pores hold natural chemical and pH gradients across thin walls, and that matters enormously, because every cell alive today runs on exactly that kind of gradient. Your mitochondria make energy by shoving protons across a membrane and letting them flow back. The alkaline-vent hypothesis, tied to Mike Russell and Nick Lane, says life didn't invent that trick. It inherited it from the rocks, borrowing a geological gradient until it could build its own. This is the metabolism-first camp: energy and chemistry got organized before genes turned up.
 Carbonate chimneys of a deep-sea alkaline hydrothermal vent
Carbonate chimneys of a deep-sea alkaline hydrothermal vent
Then there's Darwin's guess, the warm little pond, still very much in the running. On land, shallow pools go through wet-dry cycles that concentrate molecules and drive them to link up, and, crucially, ponds get ultraviolet light, which several of Sutherland's key reactions actually need. Some researchers figure the chemistry got going in sunlit surface water and only later got shoved into the deep during the chaos of the Late Heavy Bombardment. We may not have to choose. Different steps could easily have happened in different places.
Here's a thing that trips people up, though. We can actually trace the tree of life back to one ancestral population, LUCA, the Last Universal Common Ancestor that everything alive descends from. A major 2024 study in Nature Ecology & Evolution landed a jolt: LUCA lived shockingly early, around 4.2 billion years ago, in a stretch long written off as too hostile for life. And it was fancier than anyone expected.
What the 2024 study says LUCA was actually carrying
- A genome of respectable size
- An anaerobic metabolism running on hydrogen and carbon dioxide
- Enzymes that flag it as a heat-lover
- An early immune system
That last one is quietly staggering, because you only bother evolving defenses when something is already coming after you. LUCA wasn't a lonely pioneer. It lived in a crowded, competitive world, elbow to elbow with other lineages that left no surviving descendants.
But here's the humbling bit, and it's the part most people skip past. LUCA is not the origin of life. LUCA is already a fully working cell, with a genetic code, protein synthesis, the lot. It's the finished product of who-knows-how-much prior evolution. The event we're actually chasing, the spark that turned chemistry into the first thing that could copy itself, sits below LUCA, in a chapter that left no genes and, so far, no fossils. We can see the outcome. We can't yet see the moment.
What "alive" even means
This is the part I keep coming back to, because it's where the origin of life stops being a chemistry question and turns into something closer to physics and information.
From one direction, life looks thermodynamic. Ilya Prigogine showed that systems held far from equilibrium, with energy pouring through them, can spontaneously organize into ordered structures: a whirlpool, a candle flame, a convection cell. Jeremy England has pushed a modern version where matter driven by an energy source can restructure itself to dissipate that energy more efficiently, and self-copying happens to be a very good way to do exactly that. Seen this way, life doesn't fight the second law of thermodynamics. It works for it, as an unusually effective way to burn through a gradient. A star is one solution to that problem. A bacterium is another.
From a different direction, the deep question is about information. At some point a molecular system stopped merely reacting and started encoding, carrying a heritable sequence that could be copied, tweaked, and selected. The birth of the genetic code, the moment chemistry went symbolic, might be the true line between non-life and life. Which is maddening, because that's hard to even define cleanly, let alone dig out of ancient rock.
That's why one of the sharper recent ideas comes from trying to measure the boundary instead of describe it. Assembly theory, from Lee Cronin and Sara Walker, proposes you can quantify how much selection history a molecule carries by counting the steps it takes to build from basic pieces. Simple molecules form by chance all the time. Complex ones, present in real abundance, are effectively impossible to make by chance alone. The only known way to churn them out in quantity is a system that remembers how, which is to say life, or something close enough. The appeal is that this gives you a general signature that doesn't care about the substrate. Alien biology wouldn't have to resemble ours in the slightest. You'd just have to find molecules too complex to be an accident. It's an honest attempt to turn "I know it when I see it" into a number, and it doubles as a life detector you could pack onto a spacecraft.
Are we the only draft?
Which brings the whole thing to its sharpest point. Right now, life on Earth is a sample size of one. Every organism we've ever studied is related. We have exactly one data point for the most important process in the universe, and you can't draw a curve through one point.
So the endgame is to find a second genesis: life that got started independently, somewhere else. Find that, even a single microbe running a different biochemistry, and the question flips over. It would mean abiogenesis is something the cosmos does readily, given water and chemistry and time, rather than a one-off fluke. It would mean the universe is almost certainly crawling with life.
 Cassini's backlit view of Enceladus firing water-ice jets from its hidden ocean
Cassini's backlit view of Enceladus firing water-ice jets from its hidden ocean
The closest places to look are unnervingly good:
- Enceladus, a small moon of Saturn, fires plumes of water vapor straight out of a hidden ocean into space. The Cassini probe flew through them and found organics, hydrogen, and hints of hydrothermal activity on the ocean floor. Chemically, it's the alkaline-vent hypothesis you could fly a spacecraft into, and mission concepts to sample those plumes are on the table now.
- Europa, a moon of Jupiter, keeps a saltwater ocean under its ice with more liquid water than every ocean on Earth combined. NASA's Europa Clipper is already on its way.
- Titan is the most Earth-like body in the solar system in entirely the wrong way. It has a weather cycle, rivers, and seas, except they're liquid methane, with a water ocean tucked underneath. NASA's Dragonfly, a nuclear-powered drone, will fly around it and test whether life needs water at all or whether some other solvent will do.
- Mars had warm, wet, habitable conditions early on, right in the window when life was getting going on Earth. Samples are being collected now for eventual return.
- And far out among the stars, exoplanet atmospheres are being read for biosignatures. The running argument over a possible dimethyl-sulfide signal at the planet K2-18b, batted around from 2023 into 2025, is a preview of how contested, and how thrilling, that hunt is about to get.
The threshold
Here's what gets me about all of it. We are, quite literally, chemistry that woke up and started asking how it woke up. Every atom in you was forged in a star, tumbled through space, maybe rode down on something like Bennu, and got swept into a self-sustaining chemical loop that has run without a single break for four billion years, straight through to the sentence you're reading.
And we still can't tell you how the loop got started. Not really. What we've got is a pile of good candidates and no verdict. A few plausible environments. A couple of competing chemistries. That gorgeous, unsettled business with handedness. A hint from physics that life might sit closer to an expectation than an accident. And a short list of nearby worlds where the same thing might have happened a second time. The gap between "warehouse full of parts" and "a thing that copies itself and evolves" is the one that still isn't bridged, and bridging it would be one of the big ones.
What I like about the problem is that it refuses to stay in its lane. You can't crack it as just a chemist, or just an astronomer. It drags in geology and thermodynamics and information theory, and yes, particle physics, and makes them sit down together. The origin of life is the one question that quietly forces all the sciences into the same room. After seventy years of everybody being stuck, they finally seem to be talking.